Evolutieleer en natuurlijke selectie
Jan Chandesais, voorjaar 2016
Abstract
Sedert de ontdekking van de genetica groeide het inzicht dat het begrip natuurlijke selectie in essentie betekent dat de natuur genen selecteert, en sedert de bestseller van R. Dawkins aanvaardt een brede strekking binnen de wetenschap de hypothese dat eigenschappen evolueren omdat dit in het belang is van de genen die ervoor coderen. Maar een ontleding van het proces van natuurlijke selectie toont aan dat deze hypothese niet realistisch is. Natuurlijke selectie is een gebeuren dat bestaat uit deelgebeurtenissen die volgens bepaalde wetmatigheden verlopen.
Zo’n gebeuren wordt ook proces genoemd en de voorstelling ervan in een stroomdiagram geeft inzicht in de wetmatigheden. Het stroomdiagram dat het proces van natuurlijke selectie voorstelt, toont dat fitnessverandering de enige gebeurtenis is die in een wetmatig verband staat met de selectie van genen. Enkel de organismen en de grotere organische gehelen kunnen fitness realiseren en het voortbestaan van genen is daar louter een gevolg van. In die zin is Dawkins’ hypothese dat eigenschappen evolueren omdat dit in het belang is van de genen die ervoor coderen, een misvatting.
Hierbij aansluitend is ook de multilevel selection theory (E. Sober & D.S. Wilson, 1994) een misvatting. Deze theorie stelt dat het proces van natuurlijke selectie zich voordoet op drie niveaus: dat van de genen, dat van de organismen en dat van de grotere organische gehelen. De misvatting ligt erin dat de theorie geen onderscheid maakt tussen het niveau waarop de natuur kan selecteren, namelijk dat van de genen, en de niveaus waarop fitness gerealiseerd kan worden, namelijk dat van de organismen en organische gehelen.
De herkomst van fitness is deels een aangeboren aanleg en deels gedragsafhankelijk, maar voor de verdere uiteenzetting is deze nuancering onbelangrijk.
Betekenis van natuurlijke selectie
De volledige titel van Darwins ‘On the Origin of Species (1859)’ luidt: ‘Over het ontstaan van soorten door middel van natuurlijke selectie of het behoud van bevoordeelde rassen in de strijd om het leven’. Darwin had waargenomen dat het aantal nakomelingen van een soort altijd groter is dan het aantal ouders en dat na verloop van tijd de populaties toch ongeveer even groot blijven. Hij verklaarde dit door te veronderstellen dat de leden van een populatie een strijd voeren om hun bestaan (struggle for life). Deze strijd bestaat tussen groepsleden onderling, tussen groepen van eenzelfde soort en tussen de soorten. Darwin stelde ook vast dat er verschillen zijn tussen de eigenschappen van soortgenoten (variatie), dat eigenschappen worden doorgegeven aan nakomelingen en dat organismen met eigenschappen die hen een grotere fitness bezorgen, meer kans hebben om te overleven (survival of the fittest). Zijn bevindingen brachten hem tot de conclusie dat de natuur die eigenschappen selecteert die hun dragers de grootste fitness bezorgen. Dit mechanisme noemde hij natuurlijke selectie.
De genetica ontdekte dat genen coderen voor de eigenschappen van hun dragers en dat het DNA dat deze genen bevat, wordt overgedragen via voortplanting. Sedertdien groeide het inzicht dat het eigenlijke evolutieproces zich op het niveau van de genen afspeelt. Dit inzicht wordt door volgende bevindingen uit vorige eeuw duidelijk verwoordt
Francis Crick stelt in zijn centraal dogma (1958) dat de overdracht van genetische informatie naar het biologisch systeem van een organisme uitsluitend in één richting gebeurt:
The central dogma of molecular biology deals with the detailed residue-by-residue transfer of sequential information. It states that such information cannot be transferred back from protein to either protein or nucleic acid.
G.C. Williams beschrijft in zijn basiswerk (1966) de genen als onzichtbare deeltjes die het erfelijk materiaal samenstellen en de organismen als sterfelijke dragers ervan. Op blz. 24 van de e-tekst lezen we bv:
In this book I use the term gene to mean "that which segregates and recombines with appreciable frequency." Such genes are potentially immortal, in the sense of there being no physiological limit to their survival, because of their potentially reproducing fast enough to compensate for their destruction by external agents.
Op het einde van de eeuw beschrijft John Maynard Smith ‘the gene-centered view of evolution (1998)’ als volgt
If the central dogma is true, and if it is also true that nucleic acids are the only means whereby information is transmitted between generations, this has crucial implications for evolution. It would imply that all evolutionary novelty requires changes in nucleic acids, and that these changes – mutations – are essentially accidental and non-adaptive in nature. Changes elsewhere – in the egg cytoplasm, in materials transmitted through the placenta, in the mother's milk – might alter the development of the child, but, unless the changes were in nucleic acids, they would have no long-term evolutionary effects.
Als we de bevindingen van Darwin samenbrengen met de kennis uit de genetica komen we tot volgende definitie van natuurlijke selectie:
Natuurlijke selectie is een proces waarbij de natuur die genen selecteert die de fitness van organismen vergroten.
Een fitnessvergroting wordt veroorzaakt door een genetische verandering of een biotoopverandering. De geselecteerde genen komen via de voortplanting in de plaats van de tot dan toe bestaande genen.
Wetmatigheden van een natuurlijk gebeuren
Natuurlijke selectie is een natuurlijk gebeuren dat uit deelgebeurtenissen bestaat die elkaar opvolgen in een keten van oorzaak en gevolg en die samen een geheel vormen. Zo’n geheel van mekaar opvolgende gebeurtenissen wordt een proces genoemd en kan worden voorgesteld in een stroomdiagram of flowchart. Een stroomdiagram is bedacht voor de analyse/ontwikkeling van een kunstmatig proces zoals een computerprogramma, terwijl we het hier gebruiken voor de analyse/ontleding van een natuurlijk gebeuren. Het belangrijkste verschil is dat de analyse van een kunstmatig proces voorafgaat aan de ontwikkeling ervan, terwijl de analyse van een natuurlijk gebeuren dient om het proces te leren begrijpen. De belangrijkste overeenkomst is de verwerving van inzicht in de wetmatigheden van een proces. Voor het stroomdiagram van een natuurlijk gebeuren gebruiken we onderstaande symbolen.

In de opeenvolging van gebeurtenissen onderscheiden we wetmatige en onzekere verbanden. Een wetmatig verband houdt de voorspelbaarheid van een volgende gebeurtenis in. Een onzeker verband doet zich voor als een gebeurtenis oorzaak is van meerdere mogelijke gevolgen. Het voorbeeld in onderstaand stroomdiagram toont dit onderscheid.
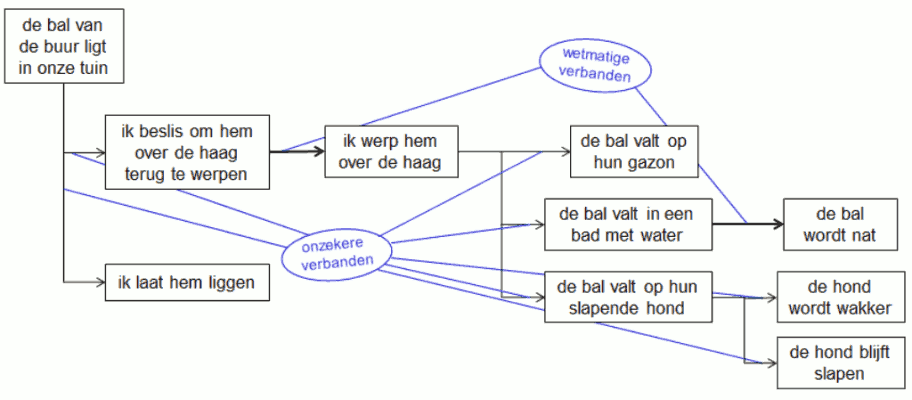
We verduidelijken nu wat we met de wetmatigheden van een natuurlijk proces bedoelen.
- We gaan ervanuit dat alle kennis over de ‘natuur in ruime zin’, waartoe de biologie en de menswetenschappen behoren, kan teruggebracht worden tot de elementaire wetten van de fysica en de scheikunde. We volgen hier de filosofie van het naturalisme die stelt dat wetenschappelijke kennis enkel kan verkregen worden over het natuurlijke en dat kennis over het bovennatuurlijke (metafysica) wetenschappelijk irrelevant is.
- Het bestaan van wetmatige verbanden tussen gebeurtenissen betekent dat ze verder geanalyseerd kunnen worden tot we terechtkomen bij de wetten van de fysica en scheikunde.
- Om een natuurlijk gebeuren te kunnen begrijpen volstaat de kennis van de deelgebeurtenissen en de aard van de verbanden ertussen, namelijk of ze onzeker of wetmatig zijn. Een stroomdiagram is een hulpmiddel om een proces tot aan deze verbanden te ontleden.
- Een wetmatig verband gaat gepaard met volledige voorspelbaarheid van de volgende gebeurtenis. Bij een verdere ontleding van zo’n verband kunnen geen onzekere verbanden gevonden worden.
- Over onzekere verbanden kan de wetenschap geen voorspelling doen, tenzij over de eventuele waarschijnlijkheid waarmee mogelijke gevolgen zich zullen voordoen. Door vordering van de wetenschap, kunnen onzekere verbanden eventueel voorspelbaar worden maar zolang dit niet het geval is, blijven ze hun aard van onzekerheid houden.
Tenslotte nog twee opmerkingen:
- De ontleding van een natuurlijk proces kan op vele manieren en hoeft niet altijd uitvoerig te zijn. Om een proces te begrijpen kan het duidelijker zijn om bepaalde verbanden niet verder te ontleden.
- We beschouwen verbanden als onzeker ofwel omdat er geen wetmatigheid mee gepaard gaat ofwel omdat de wetenschap er nog geen kennis van heeft. In beide gevallen is de veronderstelling van een wetmatigheid irrelevant.
Een voorbeeld hiervan betreft het verband tussen een genetische verandering (oorzaak) en een fitnessverandering (gevolg) dat we hieronder bespreken. Als hier een wetmatigheid mee zou gepaard gaan, kan ze wetenschappelijk (nog) niet aangetoond worden en heeft het geen betekenis om ze, zoals Dawkins in zijn bestseller doet, te veronderstellen.
Fitness is het criterium waarop de natuur selecteert
Het stroomdiagram (fig. 3) stelt het proces van natuurlijke selectie voor. De blauwe kaders geven de drie belangrijke stappen van het proces weer. Kader 1 toont de oorzaken van fitnessverandering: enerzijds genetische verandering en anderzijds verandering in de biotoop. Kader 2 toont dat fitnessverandering het criterium is waarop de natuur selecteert. Een genetische verandering beïnvloedt de fitness van organismen op twee manieren: via eigenschappen die de fitness van de gendrager zelf veranderen en via eigenschappen die de fitness van de groep, waartoe de gendrager behoort, veranderen.
Kader 3 toont dat de selectie en adaptatie van genen of allelen enkel bepaald wordt door genetische verandering en niet door een biotoopverandering.
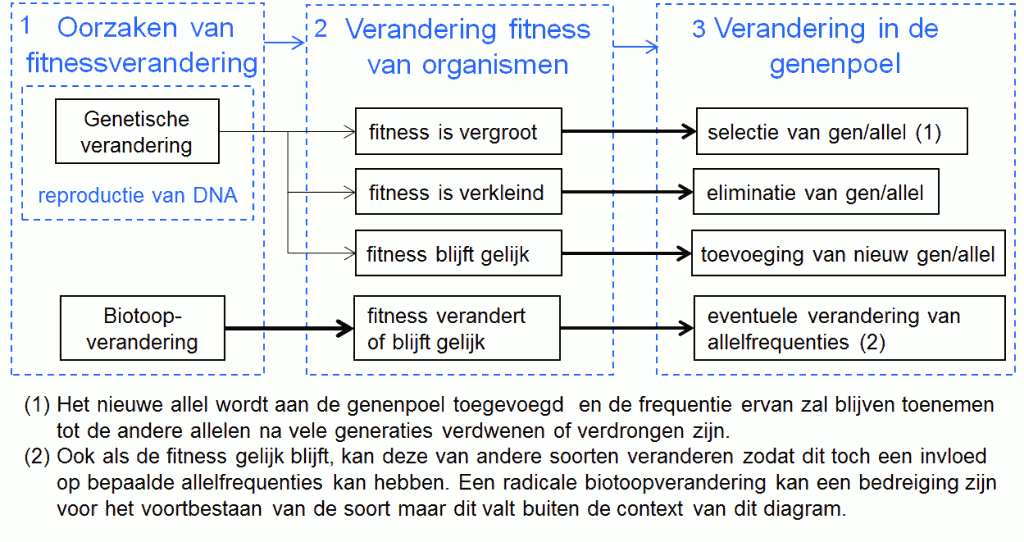
De eigenlijke evolutie van de soorten is steeds een gevolg van vernieuwing in de genenpoel, en enkel de voortplanting kan daarvan de oorzaak zijn. In de verdere tekst beperken we ons daarom tot het proces van de voortplanting (fig.4).
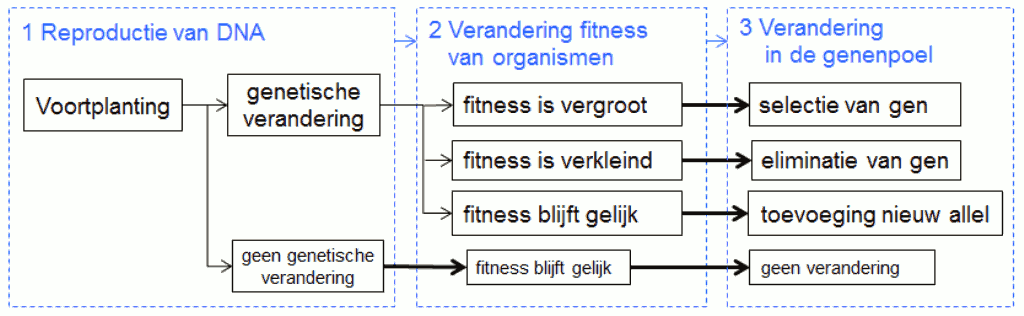
Een genetische verandering tijdens de reproductie van het DNA is een onvoorspelbare gebeurtenis, in de zin dat onze wetenschap geen kennis heeft van de oorzaken van zo’n verandering. Het verband tussen de voortplanting en een genetische verandering is dus onzeker en wordt daarom voorgesteld met een dunne verbindingspijl. Ook het verband tussen een genetische verandering en een fitnessverandering is onzeker, de wetmatigheden in het diagram blijven dus beperkt tot de verbanden tussen een fitnessverandering en veranderingen in de genenpoel.
Fitnessverandering op het niveau van de organismen is de enige gebeurtenis die op voorspelbare wijze bepaalt of genen al dan niet geselecteerd worden en er is geen terugkoppeling, namelijk het resultaat van deze selectie heeft geen invloed op gebeurtenissen die oorzaak zijn van een genetische verandering. Het ontbreken van deze terugkoppeling betekent dat de fitness van organismen het enige criterium is waarop de natuur selecteert en dat de wetmatigheid van het selectieproces zich bevindt op het niveau van de organismen en niet op dat van de genen.
Hiermee komen we niet in tegenspraak met het inzicht dat evolutie zich op het niveau van de genen afspeelt (the gene-centered view of evolution). Immers genetische verandering is een voorwaarde opdat de natuur kan selecteren (blauwe kader 1), de natuur selecteert die genen die de fitness van organismen bevorderen (kader 2) en het resultaat van deze selectie leidt tot verandering in de genenpoel (kader 3). Darwin kende de juiste mechanismen van het selectieproces niet, maar hij wist wel dat het erfelijk materiaal bepalend is voor de aangeboren eigenschappen van een organisme. Sedert vorige eeuw weten we dat erfelijke veranderingen zich voordoen op het niveau van de genen, die naargelang hun invloed op de fitness van het organisme, al dan niet geselecteerd worden.
Zelfzuchtige-genentheorie is een misvatting
Een genetische verandering is succesvol als het nieuwe gen codeert voor een eigenschap die de fitness van het organisme vergroot. Meestal is de eigenschap waarvoor een gen codeert in het belang van het organisme dat het gen draagt. In het geval van gedrag betekent dit dat de natuur zelfzuchtig gedrag selecteert.
Deze bevinding samen met de kennis van de genetica komt er op neer dat de genen voor fitness coderen en dat er verschillende niveaus van organische gehelen zijn waarop fitness kan gerealiseerd worden, namelijk de individuele organismen, de verwanten binnen een groep, de groepen, de populaties (van groepen binnen eenzelfde biotoop) en de soorten.
Het inzicht dat natuurlijke selectie zich afspeelt op het niveau van de genen en dat de natuur in essentie genen selecteert wordt ‘the gene-centered view of evolution’ genoemd, het is in overeenkomst met de theorie van Darwin en sedert midden vorige eeuw algemeen aanvaard.
Verder bouwend op dit inzicht voegde R. Dawkins er de hypothese aan toe dat het gedrag van organismen kan verklaard worden door een wetmatigheid die hij genetische zelfzucht noemt en waarmee hij bedoelt dat genen hun eigen overleving bevorderen. In zijn bestseller “The Selfish Gene (1976)” beschrijft hij het als volgt:
(e-text www.arvindguptatoys.com/arvindgupta/selfishgene-dowkins.pdf, page 29):
This book will show how both individual selfishness and individual altruism are explained by the fundamental law that I am calling gene selfishness.
(NL 2006, blz. 44):
In dit boek wil ik aantonen dat zowel individuele zelfzucht als individueel altruïsme kan worden verklaard uit een fundamentele wetmatigheid die ik genetische zelfzucht noem.
Met dit citaat maakt hij het voornemen om de wetmatigheid van genetische zelfzucht aan te tonen maar verderop in zijn boek wordt dit voornemen niet vervuld, vermoedelijk omdat deze wetmatigheid niet kan aangetoond worden. Immers de genetica leert ons dat een gen tijdens zijn individueel bestaan niks doet, het is er gewoon. Bij de celreproductie kan een gen af en toe veranderen met af en toe een positief gevolg voor de fitness van het organisme maar zo’n verandering gebeurt louter toevallig. In het besproken stroomdiagram (fig. 4) wordt dit voorgesteld door onzekere verbanden tussen een eventuele genetische verandering (kader 1) en de fitnessverandering van organismen (kader 2).
Wat ook betekent dat er geen wetmatigheid op het niveau van de genen bestaat die oorzaak is van de fitnessverandering van organismen. De enige wetmatigheid op het niveau van de genen betreft de selectie of eliminatie van genen (blauwe kader 3) en deze is louter een gevolg van wat zich op het niveau van de organismen (kader 2) afspeelt.
Bovenstaande analyse van het proces van natuurlijke selectie bevat dus geen wetmatigheid op het niveau van de genen. Maar zoals reeds gezegd kan een natuurlijk gebeuren op vele manieren door een stroomdiagram worden voorgesteld. Verdedigers van Dawkins' theorie staan dus voor een uitdaging. Als zij akkoord gaan met onze stelling dat de veronderstelde wetmatigheden van een proces enkel maar kunnen aangetoond worden door een analyse van het proces tot aan die wetmatigheden, dan is het aan hen om een analyse uit te werken die leidt tot de door Dawkins veronderstelde wetmatigheid.
Maar dan is er nog het probleem dat in Dawkins’ bestseller, naast het ontbreken van een redenering die zijn wetmatigheid aantoont, ook een selectiemechanisme ontbreekt dat deze wetmatigheid beschrijft. Het is bij gebrek aan een beschrijving van de auteur, dat wij er zelf één gemaakt hebben, namelijk:
De natuur selecteert die genen die hun eigen overlevingskansen vergroten of die hun eigen overleving bevorderen.
Tenzij men deze beschrijving kan vervangen door een betere gaan we er dus vanuit dat ze de hypothese van Dawkins juist verwoordt. Ook via deze weg komen we tot de conclusie dat de hypothese van Dawkins niet realistisch is.
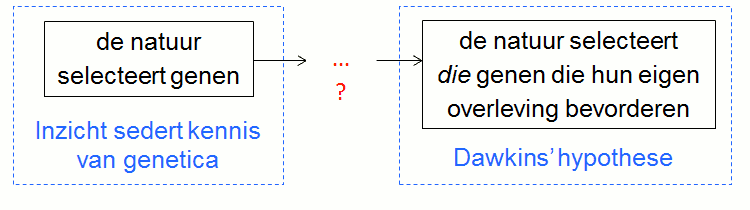
Immers, uit het inzicht ‘de natuur selecteert genen’ volgt niet dat ‘de natuur die genen selecteert die hun eigen overleving bevorderen’. Er ontbreekt namelijk een redenering die deze hypothese aantoont, wat ook betekent dat ze irrelevant is.
Dat de door Dawkins veronderstelde wetmatigheid niet bestaat, wordt tenslotte ook duidelijk als we ons de vraag stellen aan welke genen hij zelfzuchtig gedrag toeschrijft? Gaat het (1) over de genen die nog geselecteerd moeten worden of (2) over de reeds geselecteerde genen? Of bedoelt hij dat de (3) geselecteerde genen het belang dienen van genen die nog geselecteerd moeten worden. Als we deze drie mogelijkheden in detail onderzoeken, blijkt dat geen enkele tot een realistische verklaring leidt.
1. Genen hebben geen invloed op hun eigen selectie
Een gen kan haar eigen selectie niet beïnvloeden. Immers, een genetische verandering is een toevallig gebeuren en de eventuele selectie van een gen is er een louter gevolg van zonder feedback (fig.6). Een gen heeft er dus wel belang bij om geselecteerd te worden maar het kan zijn eigen selectie niet beïnvloeden.
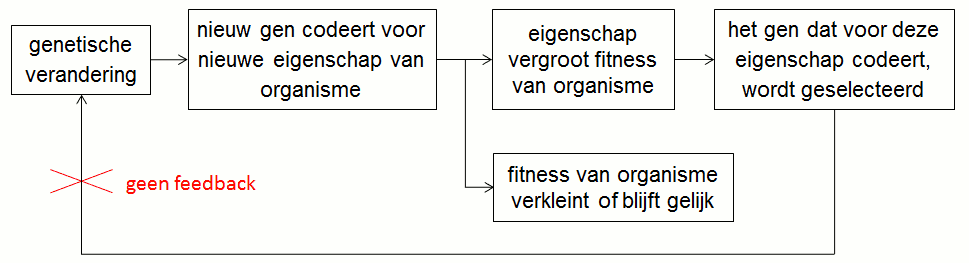
2. Reeds geselecteerde genen gedragen zich niet zelfzuchtig
Volgens de tweede mogelijkheid zijn het de reeds geselecteerde (bestaande) genen die zich zelfzuchtig gedragen, wat betekent dat ze hun eigen voortbestaan zouden bevorderen. Deze veronderstelling is niet realistisch omdat ze in tegenspraak is met de evolutieleer. Immers, volgens Darwin zijn het stapsgewijze veranderingen van het erfelijk materiaal die de essentie van evolutie uitmaken. Sedert vorige eeuw weten we dat deze veranderingen genetisch zijn: de natuur selecteert die genen die de fitness van het organisme vergroten en na verloop van tijd komen de geselecteerde genen in de plaats van de tot dan toe bestaande.
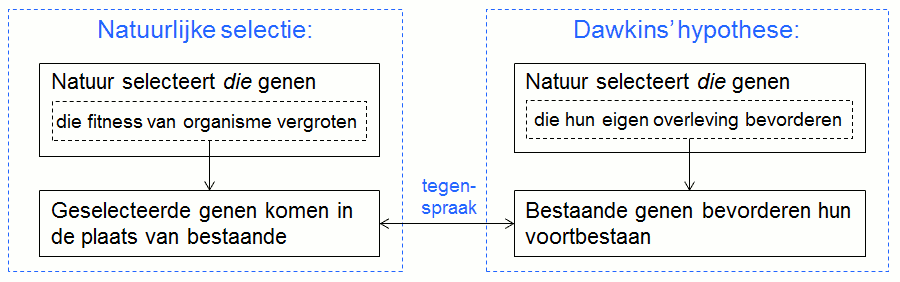
Bovenstaand diagram toont het onderscheid tussen de twee theorieën. De essentie van de evolutieleer is: door verandering van het genetisch materiaal, verhoogt de fitness van de organismen zodat de soorten veranderen of evolueren. De veronderstelling dat genen hun eigen overleving of voortbestaan bevorderen is een veronderstelling van ‘behoud van wat bestaat’ en is dus in tegenspraak met verandering of evolutie.
Het is deze tweede mogelijkheid die Dawkins vermoedelijk bedoelt. Hij benadrukt dat het eigenbelang van de genen daarin ligt dat ze zich zo snel mogelijk over zoveel mogelijk organismen willen verspreiden om hun eigen voortbestaan te verzekeren. Maar zolang het eigenbelang van genen niet wordt aangetoond, blijft dit een loze bewering. Immers het is nu eenmaal het gevolg van de selectie van een gen dat het over zoveel mogelijk organismen verspreid wordt.
3. Reeds geselecteerde genen hebben geen invloed op een genetische verandering
De resterende mogelijkheid is dat de bestaande genen het belang dienen van genen die nog geselecteerd moeten worden. We kunnen dan niet meer van eigenbelang spreken, zoals Dawkins doet, maar laat ons dit als een beeldspraak beschouwen. Niet realistisch in deze veronderstelling is dat de bestaande genen een invloed zouden hebben op een eventuele genetische verandering. Zoals reeds besproken, is dit geenszins het geval omdat een genetische verandering een louter toevallige gebeurtenis is met een onvoorspelbaar resultaat.
Multilevel selection theory bevat een redeneringsfout
Zoals gezegd was het G.C. Williams (1966) die de genen als fundamentele bouwstenen beschreef en de organismen als tijdelijke dragers. Deze opvatting wordt algemeen aanvaard en is verenigbaar met Darwins evolutieleer. Ook Dawkins bevestigt in zijn bestseller (1976) dat genen de fundamentele bouwstenen van het erfelijk materiaal zijn. De organismen zijn slechts de tijdelijke dragers terwijl de genen blijven voortbestaan. Op pag. 72 (NL 2006) lezen we bv:
Een gen mag worden beschouwd als een eenheid die voortbestaat via een groot aantal individuele lichamen. Dit is het kernpunt van het betoog dat ik in dit hoofdstuk wil ontwikkelen.
De benoeming van het gen tot ‘eenheid van selectie’ door Dawkins is dus verdedigbaar maar hij hecht er een betekenis aan die verder gaat dan deze van fundamentele bouwsteen. Op pag. 51 lezen we bv:
Ik wil hier poneren dat de fundamentele eenheid van selectie, en dus ook van het eigenbelang, noch de soort is, noch de groep, en strikt genomen zelfs niet het individu.
Dawkins stelt hier zonder bewijsvoering dat de definitie van het ‘gen als eenheid van selectie’ ook inhoudt dat het de eenheid van het eigenbelang is. Maar deze stelling volgt niet uit het inzicht dat fitness het criterium is waarop de natuur selecteert, noch uit het inzicht dat als eigenbelang de fitness van organismen verhoogt, het gen dat hiervoor codeert zal geselecteerd worden. Of het belang van een gen ‘om geselecteerd te worden’ al dan niet wordt vervuld, is gewoon een gevolg van toevallige gebeurtenissen en heeft dus niks met eigenbelang te maken.
Na het boek van Dawkins is men afgestapt van de opvatting om het gen als enige eenheid van selectie te beschouwen en tegen het eind van de eeuw wordt het begrip ‘eenheid van selectie’ gebruikt in de betekenis van ‘één der niveaus waarop de natuur selecteert’. In een artikel van E. Sober & D.S. Wilson (1994) lezen we bv:
Back to Basics. The problem of the units of selection has engaged the attention of evolutionists ever since Darwin. It concerns whether traits evolve because they benefit individual organisms' or because they are good for the group in which they occur. More recently, a third alternative has been proposed, which holds that traits evolve because they benefit the genes that code for them (Williams 1966, Dawkins 1976).
In dit citaat verwijzen de auteurs zowel naar Williams als Dawkins, alsof beide zouden beweren dat de natuur eigenschappen selecteert die in het voordeel van genen zijn, terwijl dit in het boek van Williams (1966) geenszins beweerd wordt. E. Sober & D.S. Wilson gaan er verder vanuit dat de natuur eigenschappen selecteert als ze in het voordeel zijn van de leden van een bepaalde eenheid: de genen, individuele organismen of groepen. Maar terwijl genen als voordeel hebben om geselecteerd te kunnen worden, hebben organismen het voordeel om fitness te kunnen realiseren en de auteurs gebruiken deze twee verschillende soorten voordeel in dezelfde betekenis, wat zoals zal blijken leidt tot een redeneringsfout.
Na dit uitganspunt volgt een theorie, de multilevel selection theory, die stelt dat het proces van natuurlijke selectie zich voordoet op verschillende niveaus, namelijk deze van de drie genoemde eenheden. De misvatting hiervan is dat het niveau van de genen niet in dit rijtje thuishoort. Er is immers maar één niveau van genen of bouwstenen die voor fitness coderen terwijl er meerdere niveaus van organische gehelen zijn waarop fitness nodig is in de strijd voor overleving. Deze zijn het niveau van de individuele organismen die de genen dragen, en de niveaus van grotere gehelen (groep, populatie of soort) waarbinnen de genen via voortplanting verspreid worden. Dit zijn dus ook de verschillende strijdniveaus:
- het individu is in de strijd met andere individu’s van dezelfde groep
- de groep (of populatie) is in de strijd met andere groepen van dezelfde soort
- de soort is in de strijd met andere soorten.
De eigenschappen die door evolutie ontstaan zijn slechts in het fitnessvoordeel van één van deze organische gehelen, terwijl er steeds een selectievoordeel is van de genen die de fitnessverandering realiseren. Wat het proces zelf betreft, dit kent steeds hetzelfde verloop, namelijk: een genetische verandering of biotoopverandering die oorzaak is van een fitnessverandering, die op haar beurt oorzaak is van de selectie of eliminatie van genen. Het juiste verloop van dit proces werd in detail besproken en verduidelijkt met een stroomdiagram in bovenstaande tekst.
Sedert het artikel van E. Sober & D.S. Wilson (1994) blijven aanhangers van de ‘multilevel selection theory’ ze als dusdanig verdedigen, zie bv het latere artikel van waarin de theorie verder wordt toegelicht, en zie ook de beschrijvingen van group selection en unit of selection op Wikipedia.
Los van deze misvatting, schept de ‘multilevel selection theory’ ook onduidelijkheid omdat ze er ten onrechte vanuit gaat dat de hypothese van groepselectie aanvaard is, zie het citaat ‘back to basics’ hogerop. De theorie leidt op die manier de aandacht af van de eigenlijke discussie, namelijk of fitness wel kan gerealiseerd worden op hogere niveaus dan het individuele niveau?
Referenties
P. Abbot, P. & al. (March 2011) Inclusive fitness theory and eusociality, Nature 471
Bourke, A. F. G., and N. R. Franks (1995) Social Evolution in Ants. Princeton University Press, Princeton
Darwin, Charles (1859) On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray, London – e-text www.gutenberg.org/catalog/world/readfile?fk_files=2518373
Dawkins, Richard (1976; 1989; 2006) The Selfish Gene. Oxford University Press, Oxford – e-text www.arvindguptatoys.com/arvindgupta/selfishgene-dowkins.pdf
De Waal, Frans (1996) Van nature goed. Uitgeverij Contact Amsterdam/Antwerpen
Haldane, J. B. S. (1932). The causes of evolution. London: Longman, Green
Hamilton, W. D. (1964). The genetical evolution of social behavior I. Journal of Theoretical Biology, 7, 1-16.
Hamilton, W. D. (1964). The genetical evolution of social behavior II. Journal of Theoretical Biology, 7, 17-52.
Goodall, Jane (1986) The chimpanzees of Gombe: Patterns of behavior. Harvard University Press
Koeslag, J.H. (2003). Evolution of cooperation: cooperation defeats defection in the cornfield model. J. theor. Biol. 224, 399-410
Nowak M., Tarnita C. & Wilson E. O. (August 2010). The evolution of eusociality. Nature 466, 1057–1062
Pinker, Steven (2012) The false allure of group selection – e-text edge.org/conversation/the-false-allure-of-group-selection
Smith, John Maynard (1998). Evolutionary Genetics (2nd ed.).
Sober, E., & Wilson, D. S. (1994). A critical review of philosophical work on the units of selection problem. Philosophy of Science 61 (4)
Trivers, R. L. and Hare, H. (1976). Haplodiploidy and the evolution of the social insects. Science 191:249-269.
Wynne-Edwards, V.C. (1962). Animal Dispersion in Relation to Social Behaviour. Edinburgh: Oliver & Boyd.
Williams G. C. (1966). Adaptation and Natural Selection. Princeton University Press, Princeton - e-text www.biosj.ro/hu/erdekessegek/erdekessegek_tanarok_reszerol/cikkek/WilliamsGC_Adaptation_and_Natural_Selection_1966_Princeton.pdf
Wilson, D. S., & Wilson, E. O. (2008). Evolution "for the good of the group". American Scientist, 96(5), 380-389
Wilson, E. O. (2005). Kin Selection as the Key to Altruism: its Rise and Fall. Social Research 72 (1): 159–166.
Overleg <span class="cnt_reactions">(4)</span>
Jos Vaessen
15/10/2020
Mijn samenvatting van bovenstaande is het gezegde: "Een koe in zijn gat kijken". Ofwel achteraf is alles duidelijk.
Vooraf voorspellen hoe het verspreiden van de genen en bij leven daarmee omgaan is als het gooien van een dobbelsteen. Kan een tijdje goed gaan ..........maar wat als?
Jan Chandesais
09/11/2015
Wat je opmerkingen op figuren 3/4 betreft, ik kan hier geen gewijzigde figuur toevoegen, ik zal het je een voorstel per mail bezorgen.
Wat je onderste opmerking betreft, we kunnen de evolutie van de 'mens in zijn natuurlijke situatie' als voorbeeld nemen. Deze homo is zich over de wereld gaan verspreiden zonder dat de soort veranderd is. Er zijn wel verschillen tussen bv lengte en huidskleur, die 'misschien' een gevolg zijn van hercombinatie van allelen maar die wordt geen evolutie van onze soort genoemd.
Steven Heynderickx
04/11/2015
Figuur 3/4: Proces van natuurlijke selectie
Indien een nieuw gen ontstaat door genetische verandering en het vergroot de fitness van de drager dan is er een toevoeging van dit gen (aangezien het nieuw is... het is ontstaan door genetische verandering) EN omdat het een variant is die bijdraagt tot de fitness zal na verloop van tijd dit een veelvoorkomend of veel gedragen gen zijn in de populatie. Het klopt dat de genenpoel niet groter wordt, aangezien dit gewoon een opsomming is van de bestaande varianten in de populatie, maar je mist in deze voorstelling wel de zeldzaamheid of de veel voorkomendheid van het gen.
Of de fitness nu verhoogt, verlaagt of gelijk blijft in principe komt in de poel sowieso een gen-variant bij, dus eigenlijk altijd een toevoeging. De tijd (reeksen van geslachtelijke voortplanting) en omgevingsveranderingen zullen echter laten uitwijzen of bepaalde genen in aantal toenemen, afnemen of zelfs verdwijnen.
Kader 3 heeft als titel verandering in de genenpoel, echter in de horizontale volgorde staan hier bij de onderste drie (veroorzaakt door omgevingveranderingen) niet wat er verandert met de genepoel... dat is niet consequent.
Verder lijkt het mij dat omgevingsveranderingen veranderingen veroorzaakt in de spreiding van de allelen en als gevolg mede bepaalt hoe de evolutie van een soort zal verlopen aangezien de kans dat bepaalde allelen hercombineren duidelijk zal verschillen.
Dus ik ga met boerenverstand en weinig kennis ter zake niet akkoord.
Steven Heynderickx
02/11/2015
Figuur 3/4: Proces van natuurlijke selectie
Indien een nieuw gen ontstaat door genetische verandering en het vergroot de fitness van de drager dan is er een toevoeging van dit gen (aangezien het nieuw is... het is ontstaan door genetische verandering) EN omdat het een variant is die bijdraagt tot de fitness zal na verloop van tijd dit een veelvoorkomend of veel gedragen gen zijn in de populatie. Het klopt dat de genenpoel niet groter wordt, aangezien dit gewoon een opsomming is van de bestaande varianten in de populatie, maar je mist in deze voorstelling wel de zeldzaamheid of de veel voorkomendheid van het gen.
Of de fitness nu verhoogt, verlaagt of gelijk blijft in principe komt in de poel sowieso een gen-variant bij, dus eigenlijk altijd een toevoeging. De tijd (reeksen van geslachtelijke voortplanting) en omgevingsveranderingen zullen echter laten uitwijzen of bepaalde genen in aantal toenemen, afnemen of zelfs verdwijnen.
Kader 3 heeft als titel verandering in de genenpoel, echter in de horizontale volgorde staan hier bij de onderste drie (veroorzaakt door omgevingveranderingen) niet wat er verandert met de genepoel... dat is niet consequent.
Verder lijkt het mij dat omgevingsveranderingen veranderingen veroorzaakt in de spreiding van de allelen en als gevolg mede bepaalt hoe de evolutie van een soort zal verlopen aangezien de kans dat bepaalde allelen hercombineren duidelijk zal verschillen.
Dus ik ga met boerenverstand en weinig kennis ter zake niet akkoord.
{kind=link}